|
新闻动态
|
科研进展
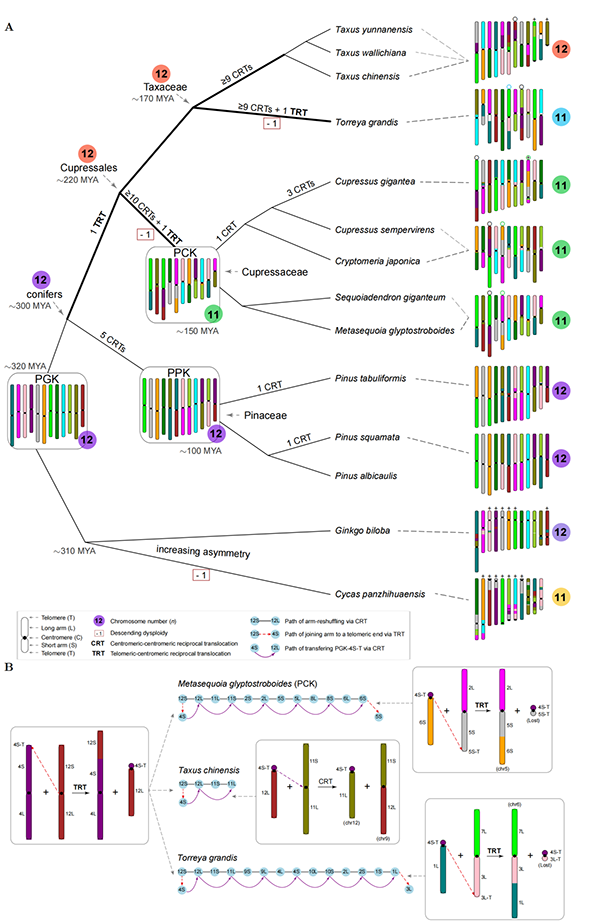
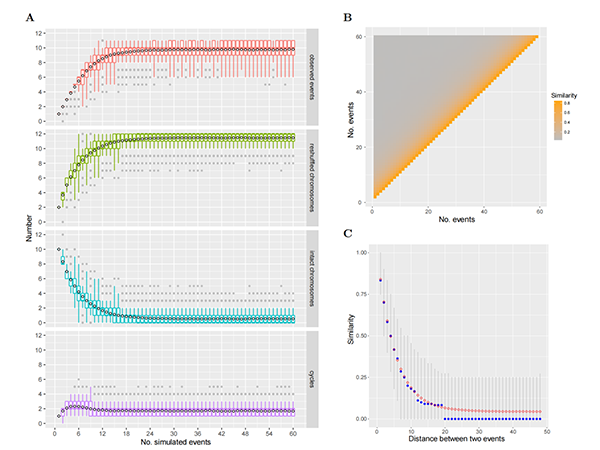
在生物演化的过程中,核型的稳定性和变异性是矛盾的两个方面。一方面,核型的稳定性对遗传信息的世代传递至关重要;另一方面,核型的变异性对物种形成和多样化也起到重要作用。不同于被子植物多样的核型,裸子植物(特别是松柏类)表现出相当高的核型一致性。这种一致性是如何形成和维持的,一直是未解之谜。中国科学院昆明植物研究所联合中国科学院植物研究所、瑞典于默奥大学研究团队,通过比较基因组分析,重建了裸子植物核型演化历史,揭示了松柏类核型演化的关键模式,为裸子植物的物种多样性演化与保护提供了新的视角。 研究团队通过对16种裸子植物染色体水平的基因组共线性分析,发现松科、柏科、红豆杉科、银杏、苏铁基因组间都显示出一对一的共线性,表明这些物种都未曾经历过谱系特有的古多倍化事件(图1),进而排除了古多倍化对裸子植物主要分支(买麻藤类除外)核型的影响。共线性模式进一步表明松柏类基因组间发生了频繁的非同源染色体之间的相互易位(reciprocal translocations, RTs),这些易位以染色体臂为单位,其断点通常都位于着丝粒附近(图2),故命名为着丝粒–着丝粒相互易位(centromeric–centromeric reciprocal translocations, CRTs)。此外,研究人员在柏目基因组中发现了另一种相互易位,其断点分别位于一条染色体的着丝粒附近和另一条染色体的端粒附近,因此命名为端粒–着丝粒相互易位(telomeric–centromeric reciprocal translocations, TRTs)。进一步的研究确认,CRTs是松柏类核型演化的最主要模式,而TRTs则在柏目植物中的递减异倍化(descending dysploidy)中发挥了关键作用。 由于CRTs反复频繁发生,解析裸子植物核型演化路径面临巨大挑战。研究团队用图论的方法(graph-based method)简化了CRTs的发生路径(图2),并进一步重建了裸子植物从共同祖先核型(proto-gymnosperm karyotype,PGK;n = 12)到现代核型(n = 11–12)的详细演化路径(图3)。重建结果显示:松科中,先发生了5次共有CRTs,之后油松中发生了一次特有CRT(chr1和chr3之间),而巧家五针松中也发生一次特有CRT(chr5和chr11之间);这些CRTs导致了7条染色体的重新组合,而剩余5条染色体则保持了祖先状态(图3A)。在柏目中情况更为复杂,几乎所有的染色体相对于PGK都发生了重新组合,模拟分析表明CRTs很可能已经饱和(图4),因此无法推断部分分支内CRTs发生的确切次数(图3A)。此外,推断柏目共同经历一次TRT事件,产生了一条只有一个祖先染色体臂的端着丝粒染色体,之后在不同分支中经历了不同的演化路径(图3B)。在柏科中,这条染色体的短臂(仅端粒)通过至少5次CRTs传递给了PGK-6S(PGK 6号染色体短臂),之后又通过一次TRT,PGK-6S的着丝粒端与PGK-5S的端粒端发生融合;形成的副产物可能是一条极短的染色体,并最终丢失,导致柏科祖先染色体数目降低到n = 11(图3B)。类似的递降异倍化也独立发生于香榧基因组(n = 11)中。在红豆杉(n = 12)中,这条端着丝粒染色体虽然也经历了一次CRT,但依旧存在至今(图3B)。总之,在柏目中,先是一次TRT改变了染色体结构和形态,之后不同谱系(柏科和香榧)独立发生的TRTs又降低了染色体数目。 研究人员还讨论了CRTs和TRTs可能的发生机制和潜在影响。CRTs频繁发生于着丝粒区域,可能是因为松柏类中普遍存在的拉布尔构型(Rabl configuration)将着丝粒聚集在一起,促进了着丝粒间的相互作用和易位。着丝粒区域大量的重复序列也能够促进非等位同源重组(non-allelic homologous recombination, NAHR)介导的相互易位。值得注意的是,反复发生的CRTs可能发生在同一着丝粒区域的不同位置。而TRTs可能偶然发生于减数分裂时期,端粒花束结构(telomere bouquet)的形成可能促进了端粒–着丝粒的互作和易位。当TRT的产物中有一条极短的染色体时,这条染色体因为不含有必需基因而可能丢失,导致递降异倍性。这与被子植物中的染色体末端融合(end-to-end fusion)类似,但被子植物异倍性的发生机制十分多样,并不局限于这种类似于TRT的机制。裸子植物超长的染色体可能限制了其他机制的发生。另外,CRTs和TRTs能够产生不同的核型、引起生殖隔离,可能促进了裸子植物的物种形成和多样化。 该研究不仅加深了我们对裸子植物核型演化的认识,而且对植物染色体演化研究具有重要参考价值。该研究成果以Convergent patterns of karyotype evolution underlying karyotype uniformity in conifers为题发表于Wiley旗下开放获取旗舰期刊Advanced Science上。中国科学院昆明植物研究所张仁纲博士研究生为论文第一作者,昆明植物所马永鹏研究员和孙卫邦研究员、瑞典于默奥大学赵伟博士和中国科学院植物研究所汪小全研究员为共同通讯作者。于默奥大学刘辉博士、昆明植物所尚鸿运博士研究生、舒恒硕士研究生(已毕业)、刘德团副研究员、杨昊硕士研究生和山东省农科院贾凯华副研究员参与了本项工作。该研究得到了国家重点研发计划(2022YFF1301700)、昆明植物所战略先导计划 (KIBXD202401)和云南省重点研发计划(202403AC100028)等项目的共同支持。
图1. 裸子植物罕见的古多倍化(WGD)事件。松科、柏科、红豆杉科、银杏、苏铁基因组间显示一对一的大规模共线性(B),表明这些物种都未曾经历过谱系特有的WGD事件(A)。同时,共线性证据也确认了前人报道的百岁兰WGD(确认为四倍化)、被子植物共有WGD(ε-WGD;确认为四倍化)和种子植物共有WGD(ζ-WGD;未知倍性);也有一定证据表明买麻藤也经历了WGD(存疑)。这些古老的WGD引发了后续的二倍化(含大量染色体重排),导致相关基因组的共线性十分破碎,相应的核型演化路径很难被追溯(A中标*处)。
图2. 松柏类基因组中的着丝粒到着丝粒的相互易位(CRTs)模式。A-C,染色体重排模式的典型案例:两种松之间可见两对非同源染色体的相互易位,且断点都发生在着丝粒(图中灰色虚线,由Hi-C信号确认)附近(A);巧家五针松和水杉(柏科)相比,所有染色体都发生了重新组合,且断点都发生在着丝粒附近(B);巧家五针松和银杏相比,7条染色体发生易位,断点在松的着丝粒附近、但偏离了银杏的着丝粒(C)。D,研究团队提出的“相继CRTs模型”,用于解释上述染色体重排模式,并用图论的方式简化重排路径。E-F,A-C对应的染色体臂重排图。
图3. 重建的裸子植物核型演化历史。A,推测的裸子植物主要分支(买麻藤类除外)的核型演化历史;B,柏目中一条端着丝粒染色体在不同分支中的演化历史,以及异倍性(dysploidy)发生途径。
图4. CRT模型的模拟。在有限的染色体数目(n = 12)下,当CRT发生次数过多时,可观察到的CRT事件趋于饱和。 |

版权所有 Copyright © 2002-2025 中国科学院昆明植物研究所,All Rights Reserved 【滇ICP备05000394号】
地址:中国云南省昆明市蓝黑路132号 邮政编码:650201
点击这里联系我们